
OS HORMÔNIOS VEGETAIS
Os Hormônios Vegetais
Uma planta precisa de diversos fatores, internos e externos, para crescer e se desenvolver, e isto inclui diferenciar-se e adquirir formas, originando uma variedade de células, tecidos e órgãos.
Como exemplos de fatores externos que afetam o crescimento e desenvolvimento de vegetais, podemos citar luz (energia solar), dióxido de carbono, água e minerais, incluindo o nitrogênio atmosférico (fixado por bactérias fixadoras e cianofíceas), temperatura, comprimento do dia e gravidade.
Os fatores internos são basicamente químicos e serão discutidos neste texto. Os principais fatores internos são os chamados hormônios vegetais ou fitormônios, substâncias químicas que atuam sobre a divisão, elongação e diferenciação celular.
Hormônios vegetais são substâncias orgânicas que desempenham uma importante função na regulação do crescimento. No geral, são substâncias que atuam ou não diretamente sobre os tecidos e órgãos que os produzem (existem hormônios que são transportados para outros locais, não atuando em seus locais de síntese), ativos em quantidades muito pequenas, produzindo respostas fisiológicas especificas (floração, crescimento, amadurecimento de frutos etc).
A palavra hormônio vem a partir do termo grego horman, que significa "excitar". Entretanto, existem hormônios inibitórios. Sendo assim, é mais conveniente considerá-los como sendo reguladores químicos.
A atuação dos reguladores químicos depende não apenas de suas composições químicas, mas também de como eles são "percebidos" pelos respectivos tecidos-alvo, de forma que um mesmo hormônio vegetal pode causar diferentes efeitos dependendo do local no qual estiver atuando (diferentes tecidos e órgãos), da concentração destes hormônios e da época de desenvolvimento de um mesmo tecido.
Os grupos de fitormônios conhecidos atualmente
Cinco grupos ou classes de hormônios vegetais (ou fitormônios) são reconhecidos:
As Auxinas
Os hormônio vegetais mais conhecidos são as auxinas, substâncias relacionadas à regulação do crescimento. Das auxinas, a mais conhecida é o AIA – ácido indolilacético.
O AIA nos vegetais não é produzido apenas em coleóptilos (Dá-se o nome de coleóptilo a primeira porção de planta que aparece à superfície do solo. Este desenvolve-se segundo a luz. Se a sua intensidade for constante, a planta irá-se desenvolver na vertical, se for iluminada lateralmente os coleóptilos irão crescer na direcção da luz, curvando-se). Sua produção também ocorre em embriões nas sementes, em tubos polínicos, e até pelas células da parede de ovários em desenvolvimento. Na planta adulta, é produzindo nas gemas apicais, principalmente as caulinares.
O transporte do AIA é polar, isto é, ocorre apenas nos locais de produção para os locais de ação por meio de células parenquimáticas especiais. O AIA age em pequeníssima quantidade, na ordem de milionésimos de mg, estimulando o crescimento.
Uma dose ótima para estimular o crescimento do caule pode inibir o crescimento da raiz.
-
A raiz e o caule de uma mesma planta reagem diferentemente ao mesmo hormônio:
A dose ótima para o crescimento da raiz é inferior à dose ótima para o crescimento do caule. A raiz, então, é mais sensível ao AIA do que caule; - A dose ótima para o crescimento do caule é inibitória para o crescimento da raiz e também inibe o crescimento das gemas laterais.
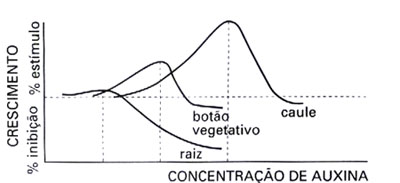
Efeito das auxinas
Na dominância apical
As auxinas atuam nos genes das células vegetais, estimulando a síntese de enzimas que promovem o amolecimento da parede celular, possibilitando a distensão das células. A forma do corpo de muitas plantas, principalmente as do grupo perene é definida pela ação hormonal. A gema apical, que atua no crescimento longitudinal do caule, produz auxina na superfície para inibie as gemas laterais, deixando-as dormentes. Eliminando-se a gema apical, o crescimento passará a ser promovido pelas gemas laterais ativadas pela ausência de auxina. O vegetal apresentará, então, forma copada: pouca altura e mais galhos.
No crescimento sob a luz
Coleóptilos submetidos à iluminação unilateral apresentaram um crescimento em direção oposta à da luz. O AIA desloca-se do lado iluminado para o não iluminado, exercendo aí o seu efeito. A curvatura do coleóptilo será tanto maior quanto maior for o tempo de iluminação, já que mais AIA acaba atingindo o lado oposto.
Se um coleóptilo for iluminado uniformemente, ele crescerá em linha reta, o mesmo acontecendo se ele for deixado no escuro.
Geotropismo
O geotropismo é um resposta dos órgãos vegetais à força da gravidade. Esta resposta resulta no crescimento da parte aérea da planta na direção oposta à força da gravidade (geotropismo negativo) e no crescimento das raízes na direção da força gravitacional (geotropismo positivo). O geotropismo no caule parece estar de acordo com a teoria de Cholodny-Went.
Quando a planta é colocada em posição horizontal, o acúmulo de auxinas na parte inferior do caule provoca um maior crescimento dessa parte, ocorrendo curvatura em uma direção oposta à força da gravidade, fazendo com que o caule se dirija para cima. Na raiz em posição horizontal ocorre um maior alongamento na parte superior comparada à inferior, provocando curvatura da raiz na direção da força gravitacional. Há pouca evidência de que ocorra uma distribuição assimétrica de AIA natural em raízes colocadas em posição horizontal.
Outros efeitos das auxinas
- A aplicação de auxinas sobre a superfície do caule promove a formação de raízes adventícias, o que é útil na propagação vegetativa por meio de estacas.
- O nível de auxinas nos tecidos do ovário sobe sensivelmente por ocasião da fecundação, promovendo o desenvolvimento do fruto.
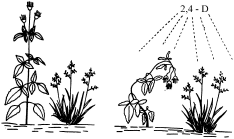
- A auxina sintética 2,4-D (ácido 2,4-diclofenoxiacético) é utilizada como herbicida e atua somente em plantas eudicotiledôneas.
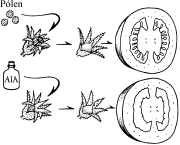
Partenocarpia
Na natureza, é comum o desenvolvimento de ovários sem que tenha havido a formação das sementes. É o caso da banana. A auxina existe na parede do ovário e também nos tubos polínicos é que garante o crescimento do fruto.
Artificialmente, é possível produzir frutos partenocárpicos por meio da aplicação de auxinas diretamente nos ovários, retirando-se previamente os estames para evitar polinização. Isso é feito para se obter uvas, melancias, e tomates sem sementes.
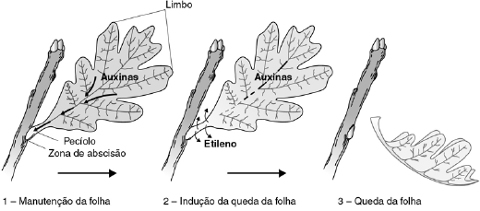
Ácido Abscísico - Abscisão Foliar
A queda das folhas de uma planta decídua pode ocorrer em resposta a sinais do meio ambiente, tais como curtos ou baixas temperaturas no outono, ou devido a condições adversas ao desenvolvimento vegetal. A folha jovem tem a capacidade de sintetizar níveis de auxinas relativamente altos; durante a senescência,a síntese de auxinas no limbo foliar diminui consideravelmente, o que promove o rompimento do pecíolo na camada de abscisão.

Durante a senescência, ao mesmo tempo que diminui o fluxo de auxinas no pecíolo, ocorre um aumento na produção de etileno na região de abscisão. A queda no nível de auxinas aparentemente torna as células da região de abscisão mais sensíveis à ação do etileno. O etileno também inibe o transporte de auxinas no pecíolo e provoca a síntese e o transporte de enzimas que atuam na parede celular (celulases) e na lamela média (pectinases). A dissolução parcial ou total da parede celular e da lamela média torna a região de abscisão enfraquecida, do ponto de vista mecânico. Basta neste momento um vento moderado para causar a quebra do feixe vascular e completar a separação da folha do restante da planta.
A abscisão de frutos é muito semelhante à abscisão foliar, somente que nos frutos e em algumas folhas ocorre, antes da abscisão, um aumento no nível de ácido abscísico. Este hormônio vegetal poderia promover a síntese de etileno e, possivelmente, a síntese das enzimas que atuam na parede celular e lamela média.
Etileno
O etileno é um hidrocarboneto insaturado, de natureza gasosa, regulador do crescimento e que atua como hormônio. Sua produção em uma planta normal ocorre praticamente em todas as células e se torna mais abundante nas flores após a polinização e nos frutos em amadurecimento. Sua síntese também se verifica em células danificadas.
Uma banana madura, colocada junto a outras verdes, acelera o amadurecimento das outras por causa do etileno que ela desprende. Por isso, os floricultores costumam armazenar frutos em câmaras onde é evitado o acúmulo de etileno no ar, retardando, assim, o amadurecimento.
Outro modo de se evitar o amadurecimento dos frutos é enriquecer o ar do armazém com gás carbônico (já que esse gás antagoniza os efeitos do etileno) ou impedir a oxigenação dos frutos (o nível baixo de oxigênio reduz a taxa de síntese de etileno).
O etileno também está envolvido com a queda – abscisão – de folhas e frutos. Esse processo começa com a redução do teor de AIA da folha, seguido pela produção do etileno. Ele estimula a síntese de celulase, enzima que digere as paredes celulósicas, na região de abscisão do pecíolo. Nessa região surge um meristema de abscisão, em que as células derivadas organizam uma cicatriz que fechará a lacuna produzida com a queda da folha ou do fruto.

As Giberelinas
A história inicial das giberelinas foi um produto exclusivo dos cientistas japoneses. Em 1926, E.Kurosawa estudava uma doença de arroz (Oryza sativa) denominada de doença das "plantinhas loucas", na qual a planta crescia rapidamente, era alta, com coloração pálida e adoentada, com tendência a cair. Kurosawa descobriu que a causa de tal doença era uma substância produzida por uma espécie de fungo, Gibberella fujikuroi, o qual parasitava as plântulas.
A giberelina foi assim denominada e isolada em 1934. As giberelinas estão presentes possivelmente em todas as plantas, por todas as suas partes e em diferentes concentrações, sendo que as mais altas concentrações estão em sementes ainda imaturas. Mais de 78 giberelinas já foram isoladas e identificadas quimicamente. O grupo mais bem estudado e o GA3 (conhecido por acido giberélico), que é também produzido pelo fungo Gibberella fujikuroi.
As giberelinas têm efeitos drásticos no alongamento dos caules e folhas de plantas intactas, através da estimulação tanto da divisão celular como do alongamento celular.
Locais de produção das giberelinas no vegetal
As giberelinas são produzidas em tecidos jovens do sistema caulinar e sementes em desenvolvimento. É incerto se sua síntese ocorre também nas raízes. Após a síntese, as giberelinas são provavelmente transportadas pelo xilema e floema.
|
Giberelinas e os mutantes anões
Giberelinas e as sementes |
 |
Aplicações práticas das giberelinas
- Giberelinas podem ser usadas na quebra de dormência de sementes de várias espécies de vegetais, acelerando a germinação uniforme de plantações. Em sementes de cevada e outras gramíneas, a giberelina produzida pelo embrião acelera a digestão em reservas nutritivas contidas no endosperma (região rica em reservas), pois estimula a produção de enzimas hidrolíticas.
- Giberelinas podem ser usadas para antecipar a produção de sementes em plantas bienais. Juntamente com as citocininas, desempenham importante papel no processo de germinação de sementes.
- Giberelinas, assim como auxinas, podem causar o desenvolvimento de frutos partenocárpicos (sem sementes), incluindo maçã, abóbora, berinjela e groselha. A maior aplicação comercial das giberelinas é na produção de uvas para a mesa. O ácido giberélico promove a produção de frutos grandes, sem sementes, soltos entre si.
- Giberelinas estimulam o florescimento de plantas de dia longo (PDL) e bienais.
Na agricultura
- Auxinas e giberelinas sintéticas: pulverizadas nas culturas, estas substâncias provocam a floração simultânea de plantações de abacaxi, evitam a queda prematura de laranjas e permitem a formação de uvas sem sementes. Aumentam ainda o tempo de armazenamento de batatas, impedindo o brotamento de suas gemas.
- Experimentos para a produção de cultura de tecidos vegetais com auxinas e citocininas em soluções nutritivivas contendo sais minerais, açúcar, vitaminas e aminoácidos. A partir disso, são produzidas grandes massas de tecidos (calos) de maçã, pêra, cenoura, batata e outros. Com estes calos, podem ser obtidas novas plantas, selecionadas e isentas de parasitas. Experimentos clássicos realizados em 1950 foram feitos para obter clones (plantas geneticamente iquais, obtidas a partir de células somáticas de um único vegetal) de cenouras por cultura de tecidos.
- Utilização de hormônios vegetais como herbicidas seletivos: alguns deles, como a 2,4 –D (ácido dicloro-fenoxiacético, uma auxina sintética) são inócuos para gramíneas como arroz, trigo, centeio, porem matam ervas daninhas de folhas largas como carrapichos, picões, dentes-de-leão.
Para outras finalidades
- Alguns hormônios sintéticos podem ser tóxicos para os animais e o homem; seu uso indiscriminado pode desencadear efeitos colaterais nocivos as comunidades e aos ecossistemas. E outra auxina sintética, a 2,4,5-T ( ácido tricloro-fenoxiacético), usado como agente desfolhante na guerra do Vietnã. Foi demonstrado que esta substância é responsável por deformações nos embriões dos mamíferos. Os efeitos perigosos da substância decorrem de sua contaminação por traços de benzodioxina, substância que se forma durante a fabricação do hormônio. Pesquisas recentes mostram que apenas cinco partes por trilhão de dioxina podem aumentar significativamente a probabilidade de ocorrência de cânceres de vários tipos.
Citocininas
Uma quarta classe de hormônios vegetais é a das citocininas, assim chamadas porque estimula a divisão celular (citocinese).
As citocininas são produzidas nas raízes e transportadas através do xilema para todas as partes da planta. Embriões e frutos também produzem as citocininas.
Funções das citocininas
O papel das citocininas no desenvolvimento das plantas tem sido estudado em culturas de tecidos. Quando um fragmento de uma planta, um pedaço de parênquima, por exemplo, é colocado em um meio de cultura contendo todos os nutrientes essenciais à sua sobrevivência as células podem crescer mas não se dividem. Se adicionarmos apenas citocinina a esse meio, nada acontece, mas se adicionarmos também auxina, as células passam a se dividir e podem se diferenciar em diversos órgãos.
|
O tipo de órgão que surge em uma cultura de tecidos vegetais depende da relação entre as quantidades de citocina e auxina adicionadas ao meio. Quando as concentrações dos dois hormônios são iguais, as células se multiplicam mas não se diferenciam, formando uma massa de células denominada calo. Se a concentração de auxina for maior que a de citocina, o calo forma raízes. Se, por outro lado, a concentração de citocina for maior do que a de auxina, o calo forma brotos.
|
As citocinas também atuam em associação com as auxinas no controle da dominância apical. Nesse caso, os dois hormônios tem efeitos antagônicos. As auxinas que descem pelo caule inibem o desenvolvimento das gemas laterais, enquanto as citocinas que vêm das raízes estimulam as gemas a se desenvolverem. Quando a gema apical é removida, cessa a ação das auxinas e as citocinas induzem o desenvolvimento das gemas laterais. Uma vez iniciado o desenvolvimento das gemas laterais não mais pode ser inibido. O fato de as gemas mais baixas do caule saírem da dormência antes das mais altas tem a ver com o fato de elas estarem mais próximas das raízes, onde são produzidas as citocinas.
As citocinas também retardam o envelhecimento das plantas. Ramos e flores cortados e colocados em água envelhecem rapidamente pela falta desse hormônio. A adição de citocina na água dos vasos faz com que as flores cortadas durem bem mais tempo. É uma prática comum no comércio de plantas pulverizar citocina sobre as flores colhidas com a finalidade de retardar o seu envelhecimento.
Senescência
|
A senescência consiste no conjunto de mudanças que provocam a deterioração e a morte da célula vegetal. Em plantas multicelulares, a senescência ocorre após a juvenilidade (crescimento vegetativo) e a maturidade (reprodução) e é rápida em plantas perenes, de acordo com o programa genético característico de cada tipo de planta. A senescência é também sensível à influência de fatores do meio ambiente tais como dias curtos, baixa luminosidade, baixas e altas temperaturas, baixos níveis de nutrientes essenciais e sais tóxicos no solo. A senescência consiste no conjunto de mudanças que provocam a deterioração e a morte da célula vegetal. Em plantas multicelulares, a senescência ocorre após a juvenilidade (crescimento vegetativo) e a maturidade (reprodução) e é rápida em plantas perenes, de acordo com o programa genético característico de cada tipo de planta. A senescência é também sensível à influência de fatores do meio ambiente tais como dias curtos, baixa luminosidade, baixas e altas temperaturas, baixos níveis de nutrientes essenciais e sais tóxicos no solo. |
 |
A senescência consiste no conjunto de mudanças que provocam a deterioração e a morte da célula vegetal. Em plantas multicelulares, a senescência ocorre após a juvenilidade (crescimento vegetativo) e a maturidade (reprodução) e é rápida em plantas perenes, de acordo com o programa genético característico de cada tipo de planta. A senescência é também sensível à influência de fatores do meio ambiente tais como dias curtos, baixa luminosidade, baixas e altas temperaturas, baixos níveis de nutrientes essenciais e sais tóxicos no solo.
A senescência e a morte podem ocorrer aproximadamente ao mesmo tempo em toda a planta, no caso de plantas anuais (milho, soja) e algumas plantas perenes (agave, bambu), que florescem uma vez e morrem logo depois, ou podem ocorrer somente na parte aérea das plantas bianuais e herbáceas perenes, nas quais as partes subterrâneas se mantêm vivas e servem como reservas para o crescimento do ano seguinte e podem ocorrer somente nas folhas e frutos de plantas lenhosas perenes. Estas plantas florescem todo ano e sua senescência total e a morte levam muitos anos.
Em plantas com senescência total, ela se dá logo depois da floração e da frutificação. A retirada de flores e de frutos adia a senescência e provoca um retorno ao rápido crescimento vegetativo característico da fase anterior à floração. Nos cereais (milho, trigo) a senescência é facilmente observada, já que acontece quase simultaneamente em milhões de plantas.
A senescência não ocorre ao acaso e quando ela é total está mais relacionada a fatores internos da planta que a fatores do meio ambiente. As mudanças que se observam nessa fase são parte de um mecanismo de transferência de nutrientes de partes da planta como a folha, para outras partes como os frutos, as sementes e o caule. Freqüentemente, a senescência das folhas e dos frutos é acompanhada de sua abscisão.
Uma causa possível da senescência da planta poderia ser a grande mobilização de nutrientes e citocininas na direção dos frutos e das sementes. A morte da parte vegetativa da planta seria a conseqüência dessa mobilização dirigida pela atividade as auxinas produzidas pelos frutos. A retirada de flores e de frutos atrasa e pode até evitar a senescência. Essa mobilização poderia também ser considerada como um efeito e não como uma causa da senescência. A teoria da mobilização não explica por que a presença de flores masculinas em plantas masculinas provoca o início da senescência enquanto a retirada dessas flores a retarda.
A aplicação de retardadores do crescimento tem como efeito a aceleração da iniciação floral e da floração de certas plantas. No abacaxizeiro, as aplicações de etileno, de compostos que liberam etileno em contato com a planta (ethephon) ou de auxinas que induzem a produção de etileno pela planta, provocam a iniciação floral e a floração. No repolho, entretanto, a floração está associada a um rápido crescimento vegetativo. Isto mostra que nem sempre a diminuição da taxa de crescimento está intimamente associada à iniciação floral, à floração e à frutificação. Na realidade, além do fato de que ela faz parte do programa genético da planta e que está sujeita, em certos casos, a fatores do meio ambiente, pouco se conhece sobre as causas profundas da senescência.